Diplodocus


Diplodocus (griego «doble viga»), o diplodoco, es un género con tres especies conocidas de dinosaurios saurópodos diplodócidos, que vivieron a finales del período Jurásico, hace aproximadamente entre 155,7 y 145 millones de años, en el Kimmeridgiense y el Titoniense, en lo que hoy es Norteamérica. Los primeros fósiles fueron descubiertos en 1877 por S. W. Williston. El nombre genérico, acuñado por Othniel Charles Marsh en 1878, es la forma en neolatín derivada del griego διπλόος (diploos) «doble» y δοκός (dokos) «viga», en referencia a la forma de los huesos a lo largo de la parte inferior de la cola. Estos huesos se pensaron exclusivos de los Diplodocus, sin embargo, han sido descubiertos en otros miembros de la familia de los diplodócidos y en saurópodos no diplodócidos como Mamenchisaurus.
Vivió en lo que es hoy el oeste de Norteamérica durante el período Jurásico Superior. El Diplodocus es uno de los dinosaurios más comunes de la parte superior de la Formación Morrison, una secuencia de sedimentos marinos y aluvionales depositados hace aproximadamente 152 millones de años. La Formación Morrison muestra un ambiente dominado por gigantescos saurópodos como Camarasaurus, Barosaurus, Apatosaurus y Brachiosaurus, además del Diplodocus.
El Diplodocus está entre los dinosaurios más fácilmente identificables, con su forma clásica, cuello y cola largos, y cuatro patas robustas. Durante muchos años, era el dinosaurio más grande conocido. Su gran tamaño pudo haber sido un impedimento para los depredadores Allosaurus y Ceratosaurus, cuyos restos se han encontrado en los mismos estratos, lo que sugiere que coexistieron con el Diplodocus.
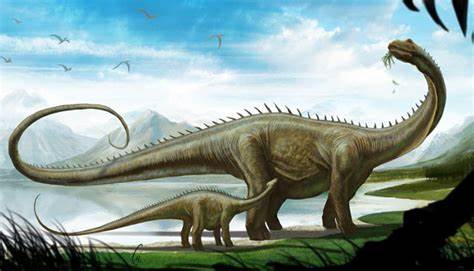
Los Diplodocus están entre los dinosaurios mejor conocidos. Fueron animales cuadrúpedos muy grandes, de cuello largo y con una extensa cola en forma de látigo. Sus miembros delanteros eran ligeramente más cortos que los traseros, lo que configura una postura predominantemente horizontal. La mecánica de estos animales de cola y cuello largos, con cuatro patas macizas, ha sido comparada con la de un puente colgante. El espécimen tipo de Diplodocus carnegii, CM 84, es el más completo hallado de su género, estimado en alrededor de 25 metros de largo y 6.5 m de alto al cuello. Para la especie de D. carnegii, conocido por restos muy bien preservados. las estimaciones de su masa se sitúan en el rango de 11,5 a 19,7 toneladas con los siguientes valores según los autores 11.5 toneladas, 12.7 toneladas, 16 toneladas,y 19,7 toneladas. Las estimaciones de masa modernas para Diplodocus carnegii tienden a estar en el rango de 10 a 16 toneladas métricas. Aunque dinosaurios como el Supersaurus eran probablemente más largos, los restos fósiles hallados de estas otras especies son sólo parciales.
Los restos parciales de D. hallorum han aumentado la longitud estimada del género, aunque no tanto como se pensó previamente. Cuando fue descrito en 1991, el descubridor, David Gillete, calculó que pudo haber medido hasta 52 metros de largo, haciéndole el dinosaurio más largo conocido, excepto los dudosos dinosaurios pobremente conocidos como Amphicoelias. Algunas estimaciones del peso se extendieron tanto como hasta 113 toneladas. La longitud estimada inicial fue posteriormente revisada a la baja a 33,5 metros y luego a 32 metros. Una revisión más reciente demuestra que las vértebras gigantes de la cola estaban colocadas realmente más adelante en la cola que lo que D. Gillete las había ubicado originalmente. Gillette había colocado originalmente las vértebras 12-19 como vértebras 20-27. El estudio demuestra que el esqueleto completo del Diplodocus en el Museo Carnegie de Historia Natural de Pittsburgh, Pensilvania, en el cual las estimaciones de Diplodocus (Seismosaurus) hallorum se basaron, incluía la decimotercera vértebra de la cola perteneciente a otro animal, lo cual redujo las estimaciones del tamaño de D. hallorum en alrededor del 30 %, quedando en aproximadamente 32 metros de largo y 25-30 toneladas.
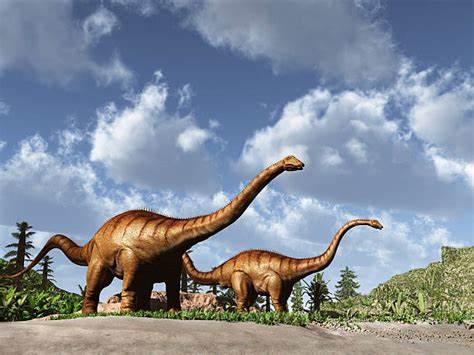
Su cuello estaba formado por al menos 15 vértebras, por lo que se cree que mantenían el cuello paralelo al suelo y que eran incapaces de elevarlo significativamente, no más de 30 grados por encima de su eje horizontal. Nunca se ha encontrado un cráneo del que se pueda decir con certeza que pertenece al Diplodocus , aunque son bien conocidos los cráneos de otros diplodócidos estrechamente relacionados con Diplodocus, como Galeamopus. Los cráneos de los diplodócidos eran muy pequeños en comparación con el tamaño de estos animales. Diplodocus tenía pequeños dientes en forma de "clavija" que apuntaban hacia adelante y solo estaban presentes en las secciones anteriores de las mandíbulas. Su caja craneana era pequeña como en otros diplodócidos. La cabeza de los Diplodocus ha sido ampliamente representada con las fosas nasales en la parte superior de la misma, debido a la posición de las aberturas nasales en el ápice del cráneo. Ha habido especulaciones acerca de si tal configuración significa que los Diplodocus tuvieron una trompa. Pero un estudio reciente afirmó que no hay evidencias paleoneuroanatómicas que respalden la hipótesis de la trompa. Se ha señalado que el nervio facial de animales como el elefante es largo, ya que inerva una trompa. Las pruebas sugieren que el nervio facial es muy pequeño en los Diplodocus. Estudios de Lawrence Witmer publicados en 2001 indicaron que, mientras que las aberturas nasales estaban arriba en la cabeza, las actuales fosas nasales carnosas estaban situadas mucho más abajo, en el hocico.
Los Diplodocus tenían una cola extremadamente larga, compuesta de más de 80 vértebras caudales, cantidad que prácticamente duplica el número con el que contaban algunos saurópodos primitivos, como el Shunosaurus con 43, y que también supera a la que tenían macronarianos contemporáneos, como el Camarasaurus con 53. Existen especulaciones acerca de que los diplódocos podrían haber utilizado su cola como defensa o para hacer ruido como un látigo. También podría haber servido como contrapeso para el cuello. La parte media de la cola tenía "dobles vigas", huesos cheurones de forma extraña que le dan a los Diplodocus su nombre. Estas pudieron haber servido para proporcionar apoyo para las vértebras, o quizás para evitar que los vasos sanguíneos se aplastasen en caso de que la pesada cola del animal chocase contra la tierra o contra algún temerario depredador. Estas "dobles vigas" también aparecen en algunos dinosaurios emparentados con los Diplodocus.
Como otros saurópodos, la mano o "pie delantero" del Diplodocus estaba altamente modificada, con los huesos del dedo y de mano dispuestos en una columna vertical, con una sección de corte con forma de herradura. El Diplodocus carecía de garras a excepción del dedo 1 de la mano, que eran inusualmente grandes comparada a la de otros saurópodos, aplanada de lado a lado, y separados de los huesos de la mano. La función de esta garra inusualmente especializada es desconocida.
Recientes descubrimientos han demostrado que los Diplodocus y otros diplodócidos pudieron haber tenido unas estrechas y puntiagudas espinas de queratina alrededor del dorso, como las de una iguana, de hasta 18 centímetros de alto en su cola y posiblemente a lo largo de la espalda y el cuello también, como en los hadrosauridos. Esta característica, radicalmente diferente de la imagen que se tenía de estos animales, ha sido incorporada en recientes reconstrucciones, como en Walking with Dinosaurs. La descripción original de las espinas informó que los especímenes en la Cantera Howe cerca de Shell, Wyoming estaban asociados con restos óseos de un diplodócido no descrito "parecido a Diplodocus y Barosaurus. Desde entonces, los especímenes de esta cantera han sido referidos a Kaatedocus siberi y Barosaurus sp., En lugar de Diplodocus.
Varias especies de Diplodocus fueron descritas entre 1878 y 1924. El primer esqueleto fue hallado en el año 1878 en Como Bluff, Wyoming, por Benjamin Mudge y Samuel Wendell Williston. Fue el paleontólogo Othniel Charles Marsh quien le puso el nombre de Diplodocus longus, "doble viga larga", ese mismo año. Desde entonces, restos de Diplodocus han sido encontrados en la Formación de Morrison, al oeste de Estados Unidos, en Colorado, en Utah, Montana y Wyoming. Los fósiles de estos dinosaurios son relativamente comunes, a excepción del cráneo que nunca se ha encontrado completo.
Los dos géneros de saurópodos de la Formación de Morrison, Diplodocus y Barosaurus, comparten huesos muy similares en sus miembros. En el pasado, muchos huesos aislados fueron automáticamente atribuidos a los Diplodocus pero pudieron, de hecho, haber pertenecido al Barosaurus.

Diplodocus longus, la especie tipo original , se conoce a partir de dos vértebras caudales fragmentarias completas y varias más de la Formación Morrison en la Cantera Felch de Colorado. Aunque varios especímenes más completos se han atribuido a D. longus, análisis detallados han sugerido que el fósil original carece de las características necesarias para permitir la comparación con otros especímenes. Por esta razón, se ha considerado un nomen dubium, que no es una situación ideal para la especie tipo de un género conocido como Diplodocus. Se está considerando una petición a la Comisión Internacional de Nomenclatura Zoológica que propone hacer que D. carnegii la nueva especie tipo.
D. lacustris es una especie dudosa, nombrada por Marsh en 1884, a partir de restos de un pequeño animal proveniente de Morrison, Colorado. En la actualidad se cree que estos huesos pertenecieron a un animal inmaduro, en vez de una especie separada. En 2015, se concluyó que el espécimen en realidad pertenecía a Camarasaurus
D. carnegii, también escrito D. carnegiei, llamado así por Andrew Carnegie, es el más conocido, principalmente debido a un esqueleto casi completo, espécimen CM 84 coleccionado por Jacob Wortman, del Museo Carnegie de Historia Natural en Pittsburgh, Pennsylvania y fue descrito y nombrado por John Bell Hatcher en 1901. Este fue considerado como especie tipo para Diplodocus por Tschopp en 2016. Aunque no es la especie del holotipo, la más conocida es Diplodocus carnegie, siendo la más famosa debido al gran número de exhibiciones que se han realizado de su esqueleto en museos de todo el mundo.
Diplodocus hayi, conocido por un esqueleto parcial y cráneo descubierto por William H. Utterback en 1902 cerca de Sheridan, Wyoming, fue descrito en 1924. En 2015, fue renombrado como el género separado Galeamopus y varios otros especímenes Diplodocus fueron referido a ese género, sin dejar conocidos calaveras de Diplodocus.
Diplodocus hallorum fue descrito por primera vez en 1991 por Gillette como Seismosaurus halli de un esqueleto parcial que comprende vértebras, pelvis y costillas, espécimen NMMNH P-3690, fue encontrado en 1979. Como el nombre específico honra a dos personas, Jim y Ruth Hall, George Olshevsky más tarde sugirió enmendar el nombre como S. hallorum, usando el genitivo plural obligatorio, Gillette luego modificó el nombre, cuyo uso fue seguido por otros, incluido Carpenter en 2006. En 2004, una presentación en la conferencia anual de la Sociedad Geológica de América argumento para que Seismosaurus sea considerado un sinónimo menor de Diplodocus. Esto fue seguido por una publicación mucho más detallada en 2006, que no solo renombró a la especie Diplodocus hallorum, sino que también apuntó que podría ser la misma que D. longus. La posición de que D. hallorum debe considerarse como un espécimen de D. longus también fue tomada por los autores de una redescripción de Supersaurus, refutando una hipótesis previa de que Seismosaurus y Supersaurus eran lo mismo. Un análisis de 2015 de las relaciones diplodócidos señaló que estas opiniones se basan en los especímenes más completos referidos de D. longus. Los autores de este análisis concluyeron que esos especímenes eran de hecho la misma especie que D. hallorum, pero que el propio D. longus era un nomen dubium.
Diplodocus es el género tipo y da su nombre a la familia Diplodocidae, a la cual pertenece. Los miembros de esta familia, aunque de gran tamaño, poseen una estructura más delgada en comparación con la de otros saurópodos, como los titanosaurianos y los braquiosáuridos. Todos están caracterizados por largos cuellos y colas y una postura horizontal, con los miembros delanteros más cortos que los traseros. Los diplodócidos dominaron a finales del Jurásico en Norteamérica y posiblemente África y parecen haber sido reemplazados por los titanosáuridos durante el Cretácico.

Una subfamilia, Diplodocinae, fue creada para incluir a los Diplodocus y a sus parientes más cercanos, incluyendo a Barosaurus. Apatosaurus, aunque contemporáneo, tiene un parentesco más lejano, pero sigue siendo considerado un diplodócido aunque no un diplodocino, ya que es un miembro de la subfamilia Apatosaurinae. Los géneros Dinheirosaurus y Tornieria han sido también identificados como parientes cercanos de Diplodocus por algunos autores.
El grupo Diplodocoidea abarca a los diplodócidos, así como los dicreosáuridos, rebaquisáuridos, Suuwassea, Amphicoelias y posiblemente Haplocanthosaurus. Anteriormente se incluía a los Nemegtosauridae, hoy incluido dentro de Lithostrotia dentro de Titanosauria. Este clado es el grupo hermano de los camarasáuridos, braquiosáuridos y titanosaurianos; los Macronaria. El conjunto de estos taxones conforman Neosauropoda, el más grande, diverso y exitoso grupo de dinosaurios sauropodomorfos.
El siguiente cladograma se basa en el análisis filogenético realizado por Whitlock en 2011, que muestra las relaciones de Diplodocus entre los otros géneros asignados a la familia Diplodocidae:
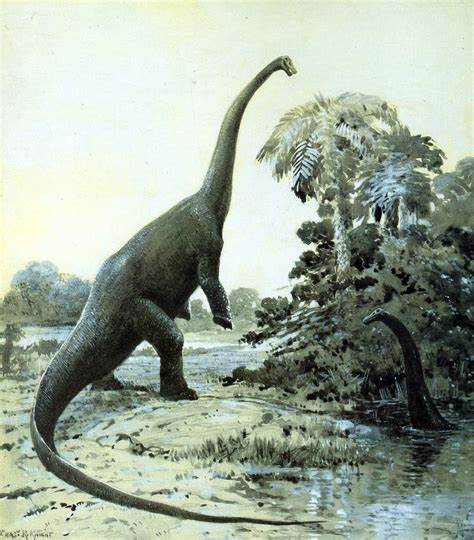
†Diplodocus
Cladograma de Diplodocidae realizado por Tschopp, Mateus y Benson en 2015, el que se realiza por especies, nótese la ausencia de D. longus ya que D. carnegii es considerado el tipo del género.
especie sin nombre

especie sin nombre

Diplodocus carnegii

Diplodocus hallorum
Debido a la gran cantidad de restos esqueléticos, Diplodocus es uno de los dinosaurios mejor estudiados. Muchos aspectos de su estilo de vida han sido objeto de diversas teorías a lo largo de los años. Las comparaciones entre los anillos esclerales de diplodócinos y las aves y reptiles modernos sugieren que pueden haber sido catemeral, activos a lo largo del día a intervalos cortos.
Marsh y luego Hatcher supusieron que el animal era acuático, debido a la posición de sus aberturas nasales en el ápice del cráneo. Comportamiento acuático similar se describió comúnmente para otros saurópodos grandes, como Brachiosaurus y Apatosaurus. Un estudio de 1951 de Kenneth A. Kermack indica que los saurópodos probablemente no podrían haber respirado por la nariz cuando el resto del cuerpo estaba sumergido, ya que la presión del agua en la pared del tórax sería demasiado grande. Desde la década de 1970, el consenso general tiene los saurópodos como animales firmemente terrestres, navegando entre árboles, helechos y arbustos.
Los científicos han debatido sobre cómo los saurópodos podían respirar con sus grandes cuerpos y largos cuellos, lo que habría incrementado la cantidad de espacio muerto . Probablemente tenían un sistema respiratorio aviar, que es más eficiente que un sistema de mamíferos y reptiles. Las reconstrucciones del cuello y el tórax de Diplodocus muestran una gran neumaticidad , que podría haber jugado un papel en la respiración como lo hace en las aves.
La representación de la postura de los Diplodocus ha cambiado considerablemente con los años. Por ejemplo, una obra clásica de 1910 de Oliver P. Hay representa a dos Diplodocus merodeando en la orilla de un río. Dichos animales cuentan con miembros abiertos hacia los lados, similares a los de los lagartos. Hay argumentó que los Diplodocus tenían un paso parecido al de un lagarto con patas sobresaliendo a ambos lados, afirmación que fue apoyada por Gustav Tornier. Sin embargo, esta hipótesis fue rebatida por William J. Holland, quien demostró que un Diplodocus con esa postura, habría necesitado un foso para hacer pasar su vientre.
Más adelante, los diplodócidos fueron retratados con frecuencia con sus cuellos elevados, lo que les permitiría comer de árboles altos, pero, recientemente, los científicos argumentaron que el corazón habría tenido problemas para mantener la presión arterial necesaria para oxigenar el cerebro. Además, estudios posteriores comprobaron que la estructura de las vértebras cervicales no pudieron permitir que el cuello se moviese tan arriba. Los estudios que analizan la morfología de los cuellos de saurópodo han concluido que la postura neutral del cuello Diplodocus era casi horizontal, en lugar de vertical, y científicos como Kent Stevens lo han usado para argumentar que los saurópodos, incluido Diplodocus, no levantaron la cabeza demasiado por encima del hombro . Un estudio de 2009 encontró que todos los tetrápodos parecen sostener la base de sus cuellos en la máxima extensión vertical posible cuando se encuentra en una postura normal y alerta y argumentó que lo mismo sería válido para los saurópodos, salvo características desconocidas y únicas que distinguen la anatomía de los tejidos blandos de sus cuellos de otros animales. El estudio encontró fallas con las suposiciones de Stevens con respecto al rango de movimiento potencial en los cuellos de saurópodos, y en base a la comparación de esqueletos con animales vivos, el estudio también argumentó que los tejidos blandos podrían tener una mayor flexibilidad de lo que sugieren los huesos. Por estas razones, argumentaron que Diplodocus habría mantenido su cuello en un ángulo más elevado que los estudios previos habían concluido.
Al igual que con el Barosaurus, el largo cuello de los Diplodocus es fuente de mucha controversia entre los científicos. En 1992, un estudio de la Universidad de Columbia sobre la estructura del cuello de un diplodócido, indicó que los cuellos tan largos habrían requerido un corazón de 1.6 toneladas. El estudio propuso que animales como éste debieron tener "corazones" auxiliares rudimentarios en sus cuellos, cuyo único propósito habría sido el de bombear la sangre al siguiente "corazón".
Comparados con los dientes de otros saurópodos, los del Diplodocus eran muy extraños. Las coronas dentales eran largas, delgadas y elípticas en sección transversal, mientras que el ápice forma un punto romo y triangular. La faceta de uso más prominente está en el ápice; sin embargo, a diferencia de los demás patrones de uso observados dentro de los saurópodos, los del Diplodocus se ubican en el lado labial, contra la mejilla, de los dientes superiores e inferiores. Esto significa que los Diplodocus y otros diplodócidos tenían un mecanismo de alimentación radicalmente diferente al de otros saurópodos. El deshojamiento de ramas usando un lado del hocico es la forma de alimentación más probable del Diplodocus, ya que explica los poco habituales patrones de desgaste de los dientes a partir del contacto entre diente y comida. Al deshojar ramas usando un lado del hocico, una fila de dientes debió usarse para arrancar el follaje del tallo, mientras que la otra debió actuar como guía y estabilizador. Gracias a que contaba con una alongada región preorbital del cráneo (delante de los ojos), podría deshojar porciones más largas de tallos en una sola acción. El movimiento hacia atrás de las mandíbulas inferiores pudo haber contribuido con dos significantes funciones en el comportamiento de alimentación, aumentar la apertura de la boca y permitir ajustes finos en las posiciones relativas de las filas de dientes para afinar el deshoje.
La flexibilidad del cuello Diplodocus se debate, pero debería haber sido capaz de navegar desde niveles bajos hasta aproximadamente 4 metros cuando está en cuatro patas. Sin embargo, los estudios han demostrado que el centro de masa de Diplodocus estaba muy cerca de la cavidad de la cadera. esto significa que Diplodocus podría volver a una postura bípeda con relativamente poco esfuerzo. También tenía la ventaja de utilizar su cola grande como un apoyo que permitiría una postura de trípode muy estable. En una postura de trípode, Diplodocus podría potencialmente aumentar su altura de alimentación hasta aproximadamente 11 metros. Con un cuello lateral y dorsoventralmente flexible y la posibilidad de utilizar su cola para erguirse apoyándose en sus miembros traseros, habilidad de «trípode», los Diplodocus habría tenido la capacidad de alimentarse de hojas en muchos niveles, bajo, intermedio y alto, hasta aproximadamente 10 metros sobre el suelo, desde los pequeños ginkgos a las colosales araucarias. El rango de movimiento del cuello también habría permitido que la cabeza pazca por debajo del nivel del cuerpo, lo que llevó a algunos científicos a especular sobre si Diplodocus pastaba en las plantas acuáticas sumergidas, de las riberas de los ríos. Este concepto de la postura de alimentación se apoya en las longitudes relativas de las extremidades anteriores y posteriores. Además, sus dientes tipo clavija se pueden haber usado para comer plantas acuáticas blanda.
Matthew Cobley y colegas en 2013 disputan lo anteriormente dicho, encontrando que los músculos grandes y el cartílago darián movimientos limitados del cuello. Afirman que los rangos de alimentación de los saurópodos como Diplodocus eran más pequeños de lo que se creía anteriormente y los animales pueden haber tenido que mover sus cuerpos para acceder mejor a las áreas donde pueden seleccionar la vegetación. Como tal, podrían haber pasado más tiempo alimentándose para satisfacer sus necesidades mínimas de energía. Las conclusiones de Cobley y colegas fueron disputadas en 2013 y 2014 por Mike Taylor, quien analizó la cantidad y posición del cartílago intervertebral para determinar la flexibilidad del cuello de Diplodocus y Apatosaurus . Taylor descubrió que el cuello del Diplodocus era muy flexible y que Cobley y colegas estaba equivocado, en afirrmar que la flexibilidad implícita en los huesos es menor.
Young y colegas en 2012 utilizaron modelos biomecánicos para examinar el rendimiento del cráneo de los diplodocinidos. Se concluyó que la propuesta de que su dentición se usara para descortezar no fue respaldada por los datos, que mostraron que bajo ese escenario, el cráneo y los dientes sufrirían tensiones extremas. Se demostró que las hipótesis de mordida de ramas y mordeduras de precisión son comportamientos de alimentación biomecánicamente plausibles. Los dientes también fueron reemplazados continuamente a lo largo de sus vidas, por lo general en menos de 35 días, como fue descubierto por Michael D'Emic y colegas. Dentro de cada alveolo dentario, se desarrollaron hasta cinco dientes de reemplazo para reemplazar el siguiente. Los estudios de los dientes también revelan que prefería una vegetación diferente a la de los otros saurópodos del Morrison, como Camarasaurus . Esto puede haber permitido mejor que las diversas especies de saurópodos existan sin competencia.
En 2010, Whitlock y colegas describieron un cráneo juvenil en el momento referido al Diplodocus, CM 11255, que difería mucho de los cráneos adultos del mismo género, su hocico no era romo y los dientes no estaban confinados al frente del hocico. Estas diferencias sugieren que los adultos y los juveniles se alimentaban de manera diferente. Tal diferencia ecológica entre adultos y juveniles no se había observado previamente en sauropodomorfos.
Al igual que otros saurópodos, a la hora de digerir los vegetales de los que se alimentaban se ayudaban de gastrolitos. La distribución de los gastrolitos hallados junto a un fósil de D. hallorum, repartidos en dos conjuntos separados, podría sugerir que estos animales tenían un sistema digestivo similar al de las aves modernas, con un buche y una molleja, y que cada grupo de gastrolitos pertenecía a cada una de estas cavidades.
Si bien el cuello largo se ha interpretado tradicionalmente como una adaptación de la alimentación, también se sugirió que el cuello sobredimensionado de Diplodocus y sus parientes podría haber sido principalmente una exhibición sexual, con cualquier otro beneficio de alimentación en segundo lugar. Un estudio de 2011 refutó esta idea en detalle.
Aunque no hay evidencia de hábitos de anidamiento de los Diplodocus, otros saurópodos como el titanosauriano Saltasaurus, han sido asociados con sitios de anidamiento. Los sitios de anidamiento de los titanosaurianos, indican que pudo haber depositado sus huevos comunalmente a lo largo de una gran área en muchos hoyos bajos, cada uno de ellos cubierto con vegetación. Esto ha generado la especulación de que los Diplodocus pudieron presentar un comportamiento similar. Otra teoría es la inclusión de las hembras en zonas de arboleda más frondosa, una vez fertilizadas, para la deposición de los huevos en un lugar "más seguro" que la desamparada planicie en la que habitualmente subsistían. El documental Walking with Dinosaurs retrató a una madre Diplodocus usando un ovipositor para poner huevos, pero fue pura especulación por parte del autor del documental. En Diplodocus y otros saurópodos, el tamaño de las nidadas y los huevos individuales eran sorprendentemente pequeños para animales tan grandes. Esto parece haber sido una adaptación a las presiones de predación, ya que los huevos grandes requerirían un mayor tiempo de incubación y, por lo tanto, estarían en mayor riesgo.
Según varios estudios de histología de huesos, los Diplodocus, al igual que otros saurópodos, crecían a un ritmo muy rápido, alcanzando la madurez sexual en tan sólo una década y continuando con su crecimiento a lo largo de toda su vida. Esta postura es radicalmente distinta de la que se atribuía a los saurópodos, los cuales se pensaba que crecían lentamente durante su vida, tomando décadas para alcanzar su madurez.
La Formación Morrison es una secuencia de sedimentos marinos y aluviales poco profundos que, según la datación radiométrica, oscila entre 156,3 millones de años en su base, y 146,8 millones de años en la parte superior, lo que lo sitúa en las etapas finales del Oxfordiense, Kimmeridgiense y Titoniense temprano del Período Jurásico Superior. Esta formación se interpreta como un entorno semiárido con distintas estaciones húmedas y secas. La cuenca de Morrison donde vivían los dinosaurios, se extendía desde Nuevo México hasta Alberta y Saskatchewan y se formó cuando los precursores de la Cordillera Frontal de las Montañas Rocosas comenzó a empujar hacia el oeste. Los depósitos de sus cuencas de drenaje orientadas al este fueron transportados por arroyos y ríos y depositados en tierras bajas pantanosas, lagos, canales fluviales y llanuras aluviales. Esta formación es similar en edad a la Formación Lourinha en Portugal y la Formación Tendaguru en Tanzania .
La Formación Morrison registra un ambiente y tiempo dominado por gigantescos dinosaurios saurópodos. Los dinosaurios conocidos del Morrison incluyen los terópodos Ceratosaurus, Koparion, Stokesosaurus, Ornitholestes, Allosaurus, Saurophaganax y Torvosaurus, los saurópodos Apatosaurus, Brontosaurus, Brachiosaurus, Camarasaurus y Diplodocus, y los ornitisquios Camptosaurus, Dryosaurus, Othnielia, Gargoyleosaurus y Stegosaurus entre otros. Diplodocus se encuentra comúnmente en los mismos sitios que Apatosaurus, Allosaurus, Camarasaurus y Stegosaurus. Allosaurus representaba del 70 al 75% de los especímenes de terópodos y se encontraba en el nivel trófico superior de la red trófica de Morrison. Muchos de los dinosaurios de la Formación Morrison son del mismo género que los que se ven en las rocas portuguesas de la Formación Lourinha, principalmente Allosaurus, Ceratosaurus, Torvosaurus y Stegosaurus o tienen una contraparte cercana, Brachiosaurus y Lusotitan, Camptosaurus y Draconyx. Otros vertebrados que compartió esta paleoambiente incluido peces con aletas radiadas, ranas, salamandras, tortugas como Dorsetochelys, esfenodontos, lagartos, terrestres y acuáticos cocodrilomorfos tales como Hoplosuchus y varias especies de pterosaurios como Harpactognathus y Mesadactylus y mamíferos primitivos docodontes, multituberculados, symmetrodontes, y triconodontes.. Conchas de bivalvos y caracoles acuáticos también son comunes. La flora del período ha sido revelada por fósiles de algas verdes, hongos, musgos, colas de caballo, cícadas, ginkgoes y varias familias de coníferas. La vegetación variaba desde los bosques de helechos arborescentes y helechos, bosques de galería, hasta las sabanas de helechos con árboles ocasionales, como la conífera Brachyphyllum de tipo Araucaria .
El Diplodocus es un dinosaurio famoso y muy representado dado que ha sido exhibido en más lugares que ningún otro saurópodo. Mucho de esto se ha debido a la abundancia de restos esqueléticos y a haber sido considerado durante mucho tiempo como el dinosaurio más largo. Al mismo tiempo, las donaciones realizadas por Andrew Carnegie, a principios del siglo XX, a potentados de todo el mundo de muchos esqueletos de yeso montados hicieron mucho para que la gente de todo el mundo se familiarice con este dinosaurio. Copias en yeso de los esqueletos de Diplodocus todavía se exhiben en muchos museos de todo el mundo, incluyendo un inusual D. hayi en el Museo de Ciencias Naturales de Houston. La serie de copias en yeso de D. carnegiei que el mecenas Andrew Garnegie y su esposa regalaron a diferentes museos e instituciones es el conjunto más famoso de esqueletos fabricados a partir del ejemplar original. Tienen como mínimo una de estas copias instituciones como el Museo de Historia Natural de Londres, el Museo Nacional de Historia Natural de Francia en París, el Museo Nacional de Ciencias Naturales en Madrid, el Museo de Historia Natural de Berlín, el Museo Senckenberg (en Fráncfort, Alemania), el Museo de Historia Natural en la Ciudad de México, el Museo Field de Historia Natural de Chicago, el Museo de La Plata en Argentina y, por supuesto, el original sigue siendo uno de los centros de atención de las colecciones del Museo Carnegie de Historia Natural en Pittsburgh. Un esqueleto montado de D. longus se encuentra en el Museo Nacional de Historia Natural de los Estados Unidos en Washington D. C., mientras que un esqueleto montado de D. hallorum (antes Seismosaurus), que puede ser el mismo que D. longus, puede ser encontrado en Museo de Nuevo México de Historia Natural y Ciencia.
Diplodocus ha sido un tema frecuente en las películas del dinosaurio, documentales y ficticias. Fue mostrado en el segundo episodio de la serie de televisión premiada de la BBC; Walking with Dinosaurs. El episodio "La Era de los Titanes" sigue la vida de un Diplodocus hace 152 millones de años, enfrentándose a ataques por parte de Ornitholestes, Stegosaurus y Allosaurus. La película animada Fantasía ofrece muchos saurópodos en el segmento de «La consagración de la primavera», uno de ellos parece ser un Diplodocus. En literatura, James A. Michener libro Centennial tiene un capítulo dedicado a Diplodocus, narrando la vida y la muerte de un individuo. Diplodocus es una figura que suele ser vista habitualmente entre los dinosaurios de juguete y entre las maquetas. Ha tenido dos modelos de juguete diferentes en la colección de juguetes Carnegie (la Carnegie Collection, véase aquí el artículo correspondiente de Wikipedia en inglés). En la película The Lost World: Jurassic Park, aparecen varios saurópodos que podrían tal vez ser identificados, algunos como Diplodocus y otros como Mamenchisaurus.
Escribe un comentario o lo que quieras sobre Diplodocus (directo, no tienes que registrarte)
Comentarios
(de más nuevos a más antiguos)
Aún no hay comentarios, ¡deja el primero!